Hafnia
Pour les articles homonymes, voir Hafnia (homonymie).
Hafnia est un genre de bacilles Gram négatifs de la famille des Hafniaceae dont il est le genre type. Son nom fait référence à Hafnia, nom médiéval de la ville de Copenhague.
| Domaine | Bacteria |
|---|---|
| Embranchement | Pseudomonadota |
| Classe | Gammaproteobacteria |
| Ordre | Enterobacterales |
| Famille | Hafniaceae |
Les Hafnia sont des bactéries de la flore digestive humaine et animale, également présentes dans l'environnement. Hafnia est souvent confondue avec Salmonella dont les caractères biochimiques sont voisins, mais l'action lytique de bactériophages spécifiques permet de les distinguer.
La principale espèce connue, Hafnia alvei, est une bactérie commensale du tractus gastro-intestinal humain et normalement non pathogène. Elle est utilisée comme ferment lactique par l'industrie laitière, et plus récemment comme un probiotique (inclus dans un produit de complément alimentaire pour la régulation de l'appétit et la perte de poids).
Historique
modifierLe genre Hafnia est l'un des 3 genres qui constituent en 2022 la famille des Hafniaceae, de l'ordre des Enterobacterales.
Bien que décrit par le biologiste danois Vagn Møller dès 1954[1], la légitimité de ce groupe a été constamment remise en question au cours des deux décennies suivantes, souvent désignées par des synonymes tels que Enterobacter alvei, Enterobacter aerogenes subsp. hafniae et Enterobacter hafniae mais, la bactérie est surtout référencée dans la littérature par son nom actuel, Hafnia alvei[2].
En 1977, une étude a conclu qu'à la fin de la fabrication du camembert, Hafnia alvei demeure l'espèce dominante. Hafnia alvei est une souche psychrotrophique, qui peut se développer à basse température, ce qui signifie qu'elle ne cesse pas de croître au cours de la phase de stockage de fromage contrairement à E. coli[3].
En 1983, la famille des Entérobactéries a été découverte dans le camembert fraîchement produit[4] : 51% des Entérobactéries ont été répertoriées comme étant des souches Hafnia alvei contre seulement 9% répertoriées comme Escherichia coli. Ces auteurs ont également montré qu'Hafnia alvei peut être trouvée à une concentration élevée dans le fromage (jusqu'à 107 UFC/g), dans le lait cru et les fromages au lait pasteurisé.
Quelques années plus tard, en 1987, Hafnia alvei a été identifiée par une équipe espagnole dans du lait cru de brebis et représentait 6,5 % du total des Enterobacteriaceae[5].
En 1987, des scientifiques américains étudiant les Brassicaceae y ont trouvé Hafnia alvei. En analysant la microflore du chou frisé fraîchement récolté, ils croyaient que l’origine d'Hafnia alvei était une contamination du sol[6].
En 2004, des scientifiques mexicains ont isolé Hafnia alvei (parmi d'autres bactéries telles que Lactobacillus acidophilus ou plantarum) de Pulque, une boisson traditionnelle fabriquée à base de maguey fermenté (également connu sous le nom d'agave)[7].
En 2014, une étude collaborative sur l'affinage du fromage, entre un laboratoire de l'INRA et une université italienne, a identifié H. alvei dans le lait cru utilisé pour faire le fromage traditionnel Caciocavallo Pugliese[8].
Hafnia alvei est également présent dans les aliments fermentés autres que les produits laitiers. Il a été identifié ces dernières années dans de nombreux plats traditionnels autour de la planète.
En 2014, Hafnia alvei a été identifié dans l'aakhone, un plat traditionnel de l'Inde du Nord à base de farine de soja fermenté[9].
Hafnia a également été identifiée dans les graines de café fermentée d'Éthiopie et dans l'Ohio aux États-Unis[10]. Des chercheurs américains ont étudié le microbiome des graines de café et identifié Hafnia alvei parmi de nombreuses autres souches et espèces, dont 13 espèces d'Enterobacteriales[11]. Cette étude a mis en évidence que la richesse du microbiome des graines était lié au processus de fermentation ainsi qu'à la qualité du café. La famille de bactéries la plus fréquemment identifiée était celle des Enterobacteriales contenant Hafnia alvei. Cette famille est souvent trouvée dans les milieux humides et riches en éléments nutritifs similaires à celles du café. Ainsi, cette bactérie pourrait jouer un rôle essentiel dans la fermentation.
Hafnia alvei a également été identifié dans le Kimchi commercial, un repas traditionnel coréen à base de chou asiatique, de radis, d'épices et de fruits de mer fermentés et salés. Les scientifiques suggèrent que la source d‘Hafnia dans ce cas est les fruits de mer fermentés[12].
Enfin, Hafnia alvei a également été isolé à partir de produits issus de la viande, en particulier du bœuf réfrigéré, en raison de la présence d' Hafnia alvei sur les bovins abattus. Il peut être trouvé aux côtés de E. Coli dans le chorizo, une saucisse espagnole semi-sèche et fermentée[7]. Les saucisses fermentées fabriquées en Espagne contiendraient également la souche Hafnia alvei, responsable de la production d'histamine, essentielle au mûrissement[13].
Souche commensale
modifierLa plupart des textes de références en microbiologie listent les mammifères, les oiseaux, les reptiles, les poissons, le sol, l'eau, les eaux usées et les aliments comme sources à partir desquelles les bactéries Hafnia alvei peuvent être récupérées.
Les voies gastro-intestinales des animaux, et en particulier des mammifères, semblent être un habitat écologique très commun pour ces bactéries. Des études de paléomicrobiologie ont permis d'identifier H. alvei provenant d'échantillons de masse intestinale et de sédiments prélevés sur des restes de mammouths vieux de 12 000 ans dans le Michigan et l'Ohio. Dans une étude portant sur 642 mammifères australiens, Gordon et FitzGibbon[14] ont trouvé qu' H. alvei était la troisième espèce entérique la plus communément identifiée, après Escherichia coli et E. cloacae.
Des Hafniae ont également été retrouvés sporadiquement dans les échantillons de fumier d'animaux de meute prélevés dans les sentiers des parcs nationaux aux États-Unis et dans 7% des grizzlis et ours noirs testés[15].
Parmi les espèces aviaires, H. alvei a été fréquemment isolé chez les oiseaux de proie, y compris les faucons, les hiboux et les vautours d'Inde; même chez les accenteurs alpins de haute altitude qui n'ont pratiquement aucun contact avec les humains, l'Hafnia a été isolée à des fréquences allant de 3% à 16%[16].
Les autres sources d' H. alvei comprennent les reptiles (serpents et scinques), les invertébrés, les insectes, les poissons et les chauves-souris.
Rôle dans la maturation du fromage
modifierSelon plusieurs publications, Hafnia alvei est présente en tant qu'espèce dominante durant la maturation du fromage au lait cru[8],[17].
Hafnia alvei est une bactérie psychrotrophique, originaire du lait cru et continue de croître dans les fromages comme le camembert. La multiplication est un facteur clé dans le processus de fermentation et de maturation du fromage.
La souche Hafnia alvei est répertoriée comme micro-organisme utilisable pour l'alimentation par l'EFFCA (European Food & Feed Cultures Association).
Depuis 1979, des études sur le fromage français ont permis d'identifier des corrélations entre la croissance d'Hafnia alvei et les paramètres chimiques lors de la fabrication du fromage. Plusieurs chercheurs ont estimé que le niveau d' H. alvei atteignait 107 CFU / g à la fin du processus de maturation et ont montré que la courbe de croissance était étroitement liée à une augmentation du pH. De même, Mounier et al. ont mesuré une population en H. alvei d'environ 109 CFU / g dans un modèle de fromage à tartiner (fromage à pâte molle)[17].
Une étude approfondie des fromages traditionnels consommés depuis des années met en évidence la présence d' Hafnia alvei dans les produits laitiers depuis plus de trente ans.
En conclusion, des taux abondants d' Hafnia alvei peuvent être trouvés dans le fromage au lait cru et jouent un grand rôle dans l'aromatisation du fromage en raison de son impact sur l'acidité et de sa capacité à produire des acides aminés libres.
L'Hafnia est soit intentionnellement ajoutée pendant le processus de fabrication du fromage, soit déjà présente comme constituant de la microflore du lait. La bactérie contribue au processus de fermentation et à la maturation des fromages. Des études métaboliques ont révélé qu' H. alvei est essentielle au processus de maturation et au développement de la saveur typique du fromage.
En raison de ces propriétés affectant l'acidité et la saveur, H. alvei est utilisé dans la fabrication de plusieurs fromages, tels que le cheddar, le gouda et le camembert, ainsi que dans le Livarot et d'autres fromages au lait cru. H. alvei est également commercialisé dans l'Union Européenne comme culture de maturation pour le camembert à fort arôme (Aroma-Prox AF 036, fourni par Bioprox SAS, France) ou comme mélange de microorganismes pour la saveur de fromage à pâte molle (Choozit Cheese cultures ARO 21 -HA LYO 10 D, fourni par Danisco, Danemark).
Une étude menée en 2013 sur l'écosystème de fromages modèles a mis en évidence le rôle d' Hafnia alvei dans l'inhibition de la croissance de la souche O26: H11 de E. coli sans modifier les concentrations de pH ou d'acide lactique. Hafnia alvei produit une petite quantité d'amines biogènes telles que la putrescine et la cadavérine, mais celles-ci n'ont pas affecté le niveau global des composés aromatiques volatils[18].
L'intérêt pour le rôle d' Hafnia alvei dans la fabrication du fromage est toujours croissant. En 2007, un projet de l'Agence Nationale de la Recherche (GRAMME) évalue les bénéfices et les risques de Hafnia alvei dans la production du fromage et étudie les autres fonctions potentielles pour élargir son application à d'autres produits alimentaires.
Culture
modifierL'hafnia croît dans des milieux contenant 2 à 5 % de chlorure de sodium, une gamme de pH de 4,9 à 8,25 et des gradients thermiques de 4 à 44 °C[19]; la température optimale pour la croissance a été rapportée à 35 °C[1].
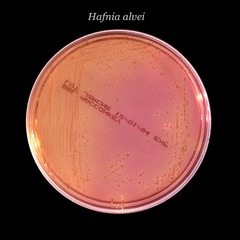
De l'avis général, presque 100 % des souches d'hafnia se développent sur les géloses MacConkey, Hektoen, éosine, bleu de méthylène et xylose-lysine-désoxycholate, qui sont tous des milieux sélectifs à modérément sélectifs[19].
Sur des milieux sélectifs les plus inhibiteurs, 25 à 60 % des souches ne poussent pas sur gélose Salmonella-Shigella (SS), tandis que 75 à 100 % des isolats sont inhibés sur un milieu vert brillant. Les souches classiques d'H. alvei sont lactose et saccharose négatives et, en tant que telles, apparaissent comme des colonies non fermentantes sur les milieux d'isolement entérique[2].
Sur les géloses modérément sélectives, elles apparaissent généralement comme de grandes colonies lisses, convexes et translucides de 2 à 3 mm de diamètre avec un bord entier; certains peuvent présenter une frontière irrégulière[2].
Biologie
modifierLipopolysaccharides
modifierL'immunochimie des lipopolysaccharides d'Hafnia est extrêmement compliquée. Tous les lipopolysaccharides d' H. alvei semblent contenir du glucose, de la glucosamine, de l'heptose et de l'acide 3-déoxyoctulosonique. Certains LPS contiennent également d'autres sucres aminés ou hydrates de carbone tels que le mannose, le galactose, la galactosamine et la mannosamine. La structure oligosaccharidique de base de certaines souches consiste en une structure hexasaccharidique identique composée de deux résidus D-glucose, de trois résidus LD-heptose et d'un résidu acide 3-déoxyoctulosonique. Il existe une grande diversité sérologique et immunologique dans ce genre, et la recherche continue dans ce domaine[20].
Biotypes
modifierEn 1969, Barbe a décrit deux biotypes d' H. alvei fondés sur la fermentation du D-arabinose et de la salicine et sur l'hydrolyse de l'esculine et de l'arbutine. L'un des défis auxquels sont confrontés les microbiologistes consiste à essayer de développer des tests biochimiques qui permettraient de distinguer facilement la plupart des souches du groupe 1 (H. alvei sensu stricto) des isolats du groupe 2 (espèces non nommées d'Hafnia). Ces deux groupes peuvent aujourd'hui être distingués les uns des autres par une série de tests. Aucun test n'étant complètement discriminatoire. Néanmoins, la mobilité à 24 heures est aujourd'hui le meilleur prédicteur du groupe (groupe 1, 9% positif, groupe 2, 100% positif)[2].
Avantages pour la santé
modifierHafnia produit une protéine appelée la Caseinolytic Protease B (ClpB) qui s'est avérée être un mimétique de l'hormone α-MSH qui est impliquée dans la satiété[21].
Il a été montré que certaines bactéries Enterobacteriacae, telles que Hafnia alvei, régulent naturellement l'appétit[22].
Plus spécifiquement, la souche Hafnia alvei HA4597 a été administrée par gavage à deux modèles d’obésité : un modèle de souris mutante obèse (souris ob/ob (en)) et un modèle de souris obèse par sur-alimentation en matières grasses. Sur ces deux modèles, la souche Hafnia alvei a montré une bonne tolérance, une réduction du gain de poids corporel et de la masse grasse dans les deux modèles d’obésité et une diminution significative de l’apport alimentaire chez les souris ob/ob[23].
Une autre étude[24] sur les souris mutantes obèses (souris ob/ob) a montré que l’administration de la souche HA4597 produisait une diminution significative (par rapport aux souris non traitées ou traitées à l’orlistat) du poids corporel, de la masse grasse et de l’apport alimentaire mais aussi chez les souris traitée une diminution de la glycémie, du cholestérol plasmatique total et de l’alanine aminotransférase (ALAT).
En 2021, les résultats d’une étude clinique de 12 semaines comparant la prise orale de la souche HA45597 et un placebo[25], ont été publiés dans la revue scientifique Nutrients. Cette étude prospective, multicentrique, double-aveugle contre placebo et randomisée chez 236 volontaires en surpoids (IMC entre 25 et 29,9 kg/m2) confirme cliniquement les données précliniques. Les deux bras ont suivi un régime hypocalorique accompagné soit du probiotique Hafnia alvei HA4597, soit d’un placebo. Le critère d'évaluation principal portant sur la perte de poids a été atteint : il y a une différence statistique significative en faveur du probiotique dans la proportion de sujets ayant perdu au moins 3% de leur poids corporel à 12 semaines (57,7% dans le groupe Hafnia contre 41,7% dans le groupe placebo, p=0,028). Parmi les critères secondaires atteints, une augmentation statistiquement significative de la sensation de rassasiement. Par ailleurs, l’effet du probiotique est également supérieur à celui du placebo dans la réduction du tour de hanche. Seul le Hafnia alvei HA4597 a eu un effet sur le taux de cholestérol et a induit une réduction de la glycémie plus importante que celle observée avec le placebo. Enfin, la souche Hafnia alvei HA4597 s’est montrée très supérieure au placebo à travers l’évaluation globale des avantages perçus tant par les médecins investigateurs que par les sujets.
La revue scientifique Nature a récemment qualifié la souche Hafnia alvei HA4597, comme probiotique de précision c’est-à-dire un probiotique dont la souche et le mécanisme d’action sont parfaitement décrits et compris[26].
TargEDys, l’entreprise française, qui a démontré que la souche Hafnia alvei HA4597 pouvait stimuler la satiété via la protéine ClpB et aider à contrôler naturellement l'appétit et à perdre du poids, a conduit les études permettant une commercialisation de ce probiotique.
Pathogénicité et sensibilité aux antimicrobiens
modifierLes informations sur la pathogénicité d'Hafnia alvei sont limitées. Bien que ce pathogène ait été retrouvé en grand nombre dans les excréments, le liquide articulaire, le sang et l'urine étaient négatifs en culture; le rôle des hafniae dans la maladie est discutable[2].
Ceci est probablement dû à la faible occurrence de cette espèce dans les maladies humaines et au fait qu'il n'y a pas de symptômes de maladie bien définis spécifiquement associés à H. alvei. Les informations actuelles concernant la pathogénicité d'Hafnia peuvent être examinées de deux points de vue; les facteurs de virulence potentiellement actifs dans les infections extra-intestinales et ceux restreints principalement à l'intestin. Une étude mentionne également que les souris dans lesquelles sont injectées par voie intrapéritonéale des hafniae n'ont pas succombé à l'infection[2].
H. alvei est un pathogène humain rare malgré l'attention accrue de la communauté médicale au cours de la dernière décennie en raison de son association possible avec la gastro-entérite.
Il n'y a pas de corrélation claire entre les données épidémiologiques, cliniques et de laboratoire et un rôle d'Hafnia alvei dans les pathologies intestinales.
Dans une étude portant sur 17 isolats d'Hafnia alvei ("Enterobacter hafniae") récupérés par la Mayo Clinic de 1968 à 1970, seuls 5 isolats (29%) ont été jugés cliniquement significatifs. Dans tous ces cas, H. alvei a été identifié comme étant un pathogène secondaire (respiratoire: 2, abcès: 3). Dans l'ensemble, l'âge moyen des personnes infectées ou colonisées par H. alvei était de 52,9 ans, avec un ratio hommes / femmes de 1: 1,1[27].
Il est intéressant de souligner que l'ATCC considère que toutes les souches d'Hafnia alvei appartiennent au niveau de biosécurité 1[28]. En outre, Richard et coll. ont identifié la présence de 108 cellules viables par gramme de fromage, ce qui suggère une consommation quotidienne de plus de 109 bactéries par jour (calcul basé sur une portion de 30 g) indiquant le bon profil d'innocuité d'Hafnia alvei dans la consommation quotidienne.
La sécurité d'Hafnia alvei chez les patients immunocompétents semble très claire et a été démontrée depuis plus de 4 décennies jusqu'à maintenant.
Stock et coll. décrivent une étude dans laquelle 76 isolats d'H. alvei ont été étudiés pour leur sensibilité à 69 antibiotiques ou médicaments. Le schéma général qui ressort de cette étude est que cette bactérie est sensible aux carbapénèmes, monobactames, chloramphénicol, quinolones, aminosides et antifolates (par exemple triméthoprime-sulfaméthoxazole) et résistante à la pénicilline, l'oxacilline et l'amoxicilline associée à l'acide clavulanique. La sensibilité aux tétracyclines et aux céphalosporines est variable[29].
Une étude espagnole, impliquant des pathogènes entériques, a également révélé que 32 souches d'H. alvei étaient universellement sensibles à toutes les quinolones (y compris la gemifloxacine et la grepafloxacine), au céfotaxime, à la gentamicine, au co-trimoxazole et à l'acide naladixique; 78% des souches de cette étude étaient sensibles à la doxycycline. Certaines souches d'H. alvei produisent à la fois des céphalosporinases inductibles de faible niveau (sensibles à la ceftazidime) et une activité céphalosporinase constitutive de haut niveau qui est résistante à la ceftazidime.
Une étude clinique testant la prise de la souche H. Alvei HA4597 chez la personne en surpoids a montré la parfaite innocuité de ce probiotique[25].
Statut réglementaire
modifierHafnia alvei est une bactérie de qualité alimentaire. Elle ne figure pas dans le catalogue Novel Food de la Commission européenne[30] mais est présente sur la liste danoise des cultures microbiennes autorisées dans les aliments[31].
Hafnia alvei fait partie de la liste publiée par l'IDF (International Dairy Federation) des cultures alimentaires microbiennes démontrant la très bonne sécurité de cette souche dans les produits alimentaires fermentés pour une utilisation mondiale[32]. Elle est aussi classée dans les produits de santé naturels par le NNHPD canadien (Natural and Non-Prescription Health Products Directorate) [33].
La souche H. alvei HA4597 est désormais commercialisée en France en tant que complément alimentaire associée à du Zinc et du Chrome sous les noms EnteroSatys Métabolisme (TargEDys[34]) et Symbiosys Satylia (Biocodex[35]), ainsi qu’avec du guarana et du Chrome sous le nom de EnteroSatys Gestion du Poids. Elle est également distribuée au Portugal (Symbiosys Satylia), en Italie (Satilia), Croatie (EnteroSatys), Turquie (Satylia), et Allemagne (SymbioLife Satylia).
Liste d'espèces
modifierSelon la LPSN (21 octobre 2022) :
- Hafnia alvei Moeller 1954
- Hafnia paralvei Huys et al. 2010
- Hafnia psychrotolerans Gu et al. 2015
Références
modifier- (en) Vagn Møller, « Distribution of amino acid decarboxylases in enterobacteriaceae », Acta Pathologica Microbiologica Scandinavica, vol. 35, no 3, , p. 259-277 (DOI 10.1111/j.1699-0463.1954.tb00869.x).
- (en) J. M. Janda et S. L. Abbott, « The Genus Hafnia : from Soup to Nuts », Clinical Microbiology Reviews, vol. 19, no 1, , p. 12-28 (ISSN 0893-8512, DOI 10.1128/cmr.19.1.12-28.2006, lire en ligne, consulté le ).
- R. Mourgues, L. Vassal, J. Auclair et G. Mocquot, « Origine et développement des bactéries coliformes dans les fromages à pâte molle », Le Lait, vol. 57, nos 563-564, , p. 131-149 (ISSN 0023-7302, DOI 10.1051/lait:1977563-5645, lire en ligne).
- J. Richard et Halima Zadi, « Inventaire de la flore bactérienne dominante des camemberts fabriqués avec du lait cru », Le Lait, vol. 63, nos 623-624, , p. 25-42 (ISSN 0023-7302, DOI 10.1051/lait:1983623-6243, lire en ligne, consulté le ).
- (en) Pilar Gaya, Margarita Medina et M. Nuntez, « Enterobacteriaceae, coliforms, faecal coliforms and salmonellas in raw ewes'milk », Journal of Applied Bacteriology, vol. 62, no 4, , p. 321-326 (ISSN 0021-8847, DOI 10.1111/j.1365-2672.1987.tb04927.x, lire en ligne, consulté le ).
- (en) S. D. Senter, J. S. Bailey et N. A. Cox, « Aerobic Microflora of Commercially Harvested, Transported and Cryogenically Processed Col lards (Brassica oleracea) », Journal of Food Science, vol. 52, no 4, , p. 1020-1021 (ISSN 0022-1147 et 1750-3841, DOI 10.1111/j.1365-2621.1987.tb14265.x, lire en ligne, consulté le ).
- (en) A Escalante, « Characterization of bacterial diversity in Pulque, a traditional Mexican alcoholic fermented beverage, as determined by 16S rDNA analysis », FEMS Microbiology Letters, vol. 235, no 2, , p. 273-279 (ISSN 0378-1097, DOI 10.1016/j.femsle.2004.04.045, lire en ligne, consulté le ).
- (en) Ilaria De Pasquale, Raffaella Di Cagno, Solange Buchin et Maria De Angelis, « Microbial Ecology Dynamics Reveal a Succession in the Core Microbiota Involved in the Ripening of Pasta Filata Caciocavallo Pugliese Cheese », Applied and Environmental Microbiology, vol. 80, no 19, , p. 6243-6255 (ISSN 0099-2240 et 1098-5336, DOI 10.1128/aem.02097-14, lire en ligne, consulté le ).
- (en) Chitta Ranjan Deb et Bendangnaro Jamir, « Nutritional Assessment and Molecular Identification of Microorganisms from Akhuni/Axone : A Soybean Based Fermented Food of Nagaland, India », Journal of Advances in Biology, vol. 11, no 1, , p. 2170-2179 (ISSN 2347-6893, DOI 10.24297/jab.v11i1.7118, lire en ligne, consulté le ).
- Tamang, J., Thapa, N., Tamang, B., Rai, A. & Chettri, R. Microorganisms in Fermented Foods and Beverages. 1–110 (2015).
- Michael Joe Vaughan, Thomas Mitchell et Brian B. McSpadden Gardener, « What's Inside That Seed We Brew? A New Approach To Mining the Coffee Microbiome », Applied and Environmental Microbiology, vol. 81, no 19, , p. 6518–6527 (ISSN 0099-2240, DOI 10.1128/aem.01933-15, lire en ligne)
- Myunghee Kim, « Isolation of Hafnia Species from Kimchi », Journal of Microbiology and Biotechnology, (ISSN 1017-7825 et 1738-8872, DOI 10.4014/jmb.0807.416, lire en ligne, consulté le )
- A Castaño, M.C Garcı́a Fontán, J.M Fresno et M.E Tornadijo, « Survival of Enterobacteriaceae during processing of Chorizo de cebolla, a Spanish fermented sausage », Food Control, vol. 13, no 2, , p. 107–115 (ISSN 0956-7135, DOI 10.1016/s0956-7135(01)00089-5, lire en ligne, consulté le )
- David M. Gordon et Frances FitzGibbon, « The distribution of enteric bacteria from Australian mammals: host and geographical effects », Microbiology, vol. 145, no 10, , p. 2663–2671 (ISSN 1350-0872 et 1465-2080, DOI 10.1099/00221287-145-10-2663, lire en ligne, consulté le )
- L. J. Goatcher, M. W. Barrett, R. N. Coleman et A. W. L. Hawley, « A study of predominant aerobic microflora of black bears (Ursus americanus) and grizzly bears (Ursus arctos) in northwestern Alberta », Canadian Journal of Microbiology, vol. 33, no 11, , p. 949–954 (ISSN 0008-4166 et 1480-3275, DOI 10.1139/m87-167, lire en ligne, consulté le )
- J. Timko et V. Kmeť, « Susceptibility of Enterobacteriaceae from the Alpine Accentor Prunella collaris », Acta Veterinaria Brno, vol. 72, no 2, , p. 285–288 (ISSN 0001-7213 et 1801-7576, DOI 10.2754/avb200372020285, lire en ligne, consulté le )
- J. Mounier, C. Monnet, T. Vallaeys et R. Arditi, « Microbial Interactions within a Cheese Microbial Community », Applied and Environmental Microbiology, vol. 74, no 1, , p. 172–181 (ISSN 0099-2240, DOI 10.1128/aem.01338-07, lire en ligne, consulté le )
- C. Delbès-Paus, S. Miszczycha, S. Ganet et S. Helinck, « Behavior of Escherichia coli O26:H11 in the presence of Hafnia alvei in a model cheese ecosystem », International Journal of Food Microbiology, vol. 160, no 3, , p. 212–218 (ISSN 0168-1605, DOI 10.1016/j.ijfoodmicro.2012.10.019, lire en ligne, consulté le )
- S. GREIPSSON et F. G. PRIEST, « Numerical Taxonomy of Hafnia alvei », International Journal of Systematic Bacteriology, vol. 33, no 3, , p. 470–475 (ISSN 0020-7713 et 1465-2102, DOI 10.1099/00207713-33-3-470, lire en ligne, consulté le )
- Ewa Katzenellenbogen, Nina A. Kocharova, George V. Zatonsky et Alexander S. Shashkov, « Structure of the O-polysaccharide of Hafnia alvei strain PCM 1189 that has hexa- to octasaccharide repeating units owing to incomplete glucosylation », Carbohydrate Research, vol. 340, no 2, , p. 263–270 (ISSN 0008-6215, DOI 10.1016/j.carres.2004.11.009, lire en ligne, consulté le )
- N Tennoune, P Chan, J Breton et R Legrand, « Bacterial ClpB heat-shock protein, an antigen-mimetic of the anorexigenic peptide α-MSH, at the origin of eating disorders », Translational Psychiatry, vol. 4, no 10, , e458–e458 (ISSN 2158-3188, DOI 10.1038/tp.2014.98, lire en ligne, consulté le )
- Sergueï O. Fetissov, « Role of the gut microbiota in host appetite control: bacterial growth to animal feeding behaviour », Nature Reviews Endocrinology, vol. 13, no 1, , p. 11–25 (ISSN 1759-5029 et 1759-5037, DOI 10.1038/nrendo.2016.150, lire en ligne, consulté le )
- Romain Legrand, Nicolas Lucas, Manon Dominique et Saida Azhar, « Commensal Hafnia alvei strain reduces food intake and fat mass in obese mice—a new potential probiotic for appetite and body weight management », International Journal of Obesity, vol. 44, no 5, , p. 1041–1051 (ISSN 0307-0565 et 1476-5497, DOI 10.1038/s41366-019-0515-9, lire en ligne, consulté le )
- Nicolas Lucas, Romain Legrand, Camille Deroissart et Manon Dominique, « Hafnia alvei HA4597 Strain Reduces Food Intake and Body Weight Gain and Improves Body Composition, Glucose, and Lipid Metabolism in a Mouse Model of Hyperphagic Obesity », Microorganisms, vol. 8, no 1, , p. 35 (ISSN 2076-2607, DOI 10.3390/microorganisms8010035, lire en ligne, consulté le )
- Déchelotte P, Breton J, Trotin-Picolo C, et al. The probiotic strain hafnia alvei HA4597® improves weight loss in overweight subjects under moderate hypocaloric diet: A proof-of-concept, multicenter randomized, double-blind placebo-controlled study. Nutrients. 2021;13:1902. https://doi.org/10.3390/nu13061902
- J. A. Washington, R. J. Birk et R. E. Ritts, « Bacteriologic and Epidemiologic Characteristics of Enterobacter hafniae and Enterobacter liquefaciens », Journal of Infectious Diseases, vol. 124, no 4, , p. 379–386 (ISSN 0022-1899 et 1537-6613, DOI 10.1093/infdis/124.4.379, lire en ligne, consulté le )
- (en) « Biosafety Level », sur www.lgcstandards-atcc.org (consulté le )
- Ingo Stock, Motiur Rahman, Kimberley Jane Sherwood et Bernd Wiedemann, « Natural antimicrobial susceptibility patterns and biochemical identification of Escherichia albertii and Hafnia alvei strains », Diagnostic Microbiology and Infectious Disease, vol. 51, no 3, , p. 151–163 (ISSN 0732-8893, DOI 10.1016/j.diagmicrobio.2004.10.008, lire en ligne, consulté le )
- https://ec.europa.eu/food/safety/novel_food/catalogue_en
- Inge Larsen, Charlotte Kristiane Hjulsager, Anders Holm et John Elmerdahl Olsen, « A randomised clinical trial on the efficacy of oxytetracycline dose through water medication of nursery pigs on diarrhoea, faecal shedding of Lawsonia intracellularis and average daily weight gain », Preventive Veterinary Medicine, vol. 123, , p. 52–59 (ISSN 0167-5877, DOI 10.1016/j.prevetmed.2015.12.004, lire en ligne, consulté le )
- Salvatore Coppola, Giuseppe Blaiotta et Danilo Ercolini, « Dairy Products », dans Molecular Techniques in the Microbial Ecology of Fermented Foods, Springer New York (ISBN 9780387745190, lire en ligne), p. 31–90
- « Canadian Natural and Non-Prescription Health Products Directorate »
- https://www.targedys.com/fr/
- https://www.biocodex.fr/fr/
Liens externes
modifier- (en) Référence Catalogue of Life : Hafnia Moller, 1954 (consulté le )
- (en) Référence NCBI : Hafnia (taxons inclus)
- « Enterosatys Complément alimentaire à base d’Hafnia alvei 4597 », sur www.enterosatys.fr (consulté le )