Conocephalum
Conocephalum est un genre d'hépatiques thalloïdes complexes de l'ordre des Marchantiales et est le seul genre existant de la famille des Conocephalaceae[1],[2]. Certaines espèces de Conocephalum sont attribuées au complexe Conocephalum conicum, qui comprend plusieurs espèces cryptiques[1]. Les espèces de Conocephalum sont de grandes hépatiques avec des motifs distincts sur le thalle supérieur, donnant l'apparence d'une peau de serpent[3]. L'espèce Conocephalum conicum doit son nom à ses structures reproductrices en forme de cône, appelées archégoniophores[4].
_3102.JPG)
| Règne | Plantae |
|---|---|
| Division | Marchantiophyta |
| Classe | Marchantiopsida |
| Sous-classe | Marchantiidae |
| Ordre | Marchantiales |
Müll.Frib. ex Grolle
Les espèces de Conocephalum sont relativement communes[5] et largement réparties en Amérique du Nord, en Europe et en Asie de l'Est[6]. Conocephalum se rencontre souvent dans des habitats humides et ombragés[1],[5],[7],[8] et se trouve également dans des forêts ouvertes, des bancs de sable, des rochers et des falaises humides et des sols humides[4]. Les espèces de Conocephalum sont également souvent associées à des substrats calcaires[7].
Conocephalum a un thalle relativement grand[4],[5] avec une ramification irrégulière[4]. Les plantes poussent en chevauchant les lobes, créant souvent de grands tapis[5]. En ce qui concerne la reproduction, les espèces de Conocephalum sont dioïques[9]. Les espèces de Conocephalum produisent différents terpènes et composés aromatiques[10],[11]. Une variation considérable des espèces a été identifiée en fonction de la composition chimique[12] et différentes espèces ont été identifiées en fonction de leurs composés uniques. Un alcool sesquiterpénique unique connu sous le nom de conocephalenol a été identifié et extrait de C. conicum[10],[11].
Classification et taxonomie
modifierCertaines espèces de Conocephalum sont placées dans le complexe Conocephalum conicum, qui comprend plusieurs espèces cryptiques[1]. Par conséquent, il a été difficile d'identifier le nombre exact d'espèces dans ce genre[13]. Une espèce cryptique fait référence à une espèce qui présente une différence génétique mais qui ne présente pas de différences morphologiques. Chez les hépatiques, on suggère que les espèces cryptiques sont liées à la fois à la disjonction géographique et à la biologie de la reproduction en combinaison avec l'isolement et la différenciation de l'habitat[12].
La recherche moléculaire a indiqué que Conocephalum comprend un complexe de six espèces cryptiques (A, C, F, J, L et S)[12],[14]. En 2005, l'espèce cryptique S de C. conicum a été décrite comme une espèce distincte, C. salebrosum[1],[7],[12]. Conocephalum salebrosum a une distribution plus large et est présent en Amérique du Nord, contrairement à C. conicum[1],[3],[7]. Des études plus récentes du complexe Conocephalum conicum au Japon et à Taiwan ont permis d'identifier trois nouvelles espèces au sein de Conocephalum, C. orientalis, C. purpureorubum et C. toyotae, qui étaient auparavant décrites comme C. conicum J, F et R respectivement[2].
Espèces
modifier- Complexe Conocephalum conicum – comprend plusieurs espèces cryptiques[1],[2] :
- Conocephalum supradecompositum
Répartition
modifier
Les espèces de Conocephalum sont réparties dans toute l'Amérique du Nord, l'Europe et l'Asie de l'Est[6],[9],[13]. Conocephalum salebrosum présente la répartition la plus large et se trouve dans toute l'Amérique du Nord[5],[7],[13], l'Europe et l'Asie[15]. En Amérique du Nord, C. salebrosum est présent dans tout le Canada et dans certaines parties des États-Unis et a également été signalé en Russie[7]. Contrairement à C. salebrosum, C. conicum est présent dans toute l'Europe[9],[13] et a été signalé en Norvège, en Finlande, en Grande-Bretagne, en Irlande, en Belgique, en France, en Allemagne, en République tchèque, en Autriche, en Hongrie, en Roumanie, en Italie, au Portugal, en Espagne, en Croatie, en Bulgarie, en Grèce, en Ukraine, en Pologne et en Russie[16].
L'espèce C. supradecompositum a une répartition plus restreinte et se trouve principalement en Chine et au Japon[17],[18]. En ce qui concerne les espèces de Conocephalum les plus récemment décrites, C. purpureorubum a été observé au Japon, en Chine, à Taiwan et en Corée du Sud et C. orientalis a été trouvé au Japon et à Taiwan[2],[9].
Habitat
modifierLes espèces de Conocephalum se rencontrent souvent dans des habitats humides et ombragés[1],[5],[7],[8]. Les espèces de Conocephalum poussent également dans des microhabitats spécialisés à proximité d'eaux courantes et stagnantes[15]. Conocephalum conicum se rencontre souvent dans des forêts ouvertes, des bancs de sable, des rochers et des falaises humides et des sols humides[4]. C. conicum et C. salebrosum sont tous deux fortement associés aux substrats calcaires[7]. Il a également été suggéré que C. salebrosum est probablement plus tolérant à la dessiccation que C. conicum[7],[8],[9],[15],[19].
Morphologie
modifier_6046.JPG)
_7776.JPG)
Conocephalum conicum et C. salebrosum partagent certaines similitudes dans leurs caractéristiques morphologiques, en plus d'avoir leurs propres traits uniques qui aident à distinguer les deux espèces[19].
_7806.JPG)
Gamétophyte
modifierLa structure végétative de Conocephalum est un thalle qui a l'apparence d'un corps aplati de tissu végétal[5],[20]. Le thalle est irrégulièrement ramifié[4] et relativement grand, atteignant une longueur d'environ 20 à 24 cm[4],[5]. Contrairement à C. conicum et C. salebrosum, le thalle de C. supradecompositum est relativement petit, mesurant 2 à 3 cm de long[18]. Le thalle se développe en développant des lobes[20] qui se fanent à mesure que la plante mûrit[21]. Les plantes de C. salebrosum poussent souvent en chevauchant les lobes, créant parfois de grands tapis[5].
Les espèces de Conocephalum ont un thalle d'apparence terne, comme C. salebrosum, ou nettement brillant, comme C. conicum[1],[9]. La surface supérieure du thalle présente des contours hexagonaux caractéristiques formés de rainures peu profondes autour de chaque chambre à air[4]. Le tissu photosynthétique et les chloroplastes sont situés dans les chambres à air[22]. Au milieu de chaque chambre à air se trouve un pore cerclé de blanc[4],[21]. Les parois supérieures des grandes chambres à air sont souvent visibles à la surface du thalle[21]. Le pore de la chambre à air reste ouvert, contrairement aux stomates des plantes vasculaires où les pores peuvent s'ouvrir et se fermer[5].
La face inférieure du thalle comporte à la fois des rhizoïdes et des écailles[4]. Les écailles sont de couleur violette et sont disposées au milieu de la face inférieure du thalle[4]. Des rhizoïdes sont également présents sur la face inférieure du thalle. Il existe deux types de rhizoïdes, les rhizoïdes longs et lisses et les rhizoïdes courts et pointus[4],[20],[23]. On pense que les rhizoïdes courts jouent un rôle dans l'absorption de l'eau et des nutriments. En revanche, les rhizoïdes plus longs aident à ancrer le thalle au substrat sous-jacent. Les rhizoïdes sont unicellulaires, contrairement aux rhizoïdes multicellulaires que l'on trouve dans les mousses[20].
Corps huileux complexes
modifierLes cellules des hépatiques contiennent souvent des corps huileux complexes[12],[20],[23],[24]. Les corps huileux sont des organites intracellulaires délimités par une seule membrane[22],[25]. On sait que les corps huileux contiennent une variété de composés phytochimiques uniques, tels que des terpènes et des flavonoïdes[24]. La fonction des corps huileux est encore mal comprise[22],[23],[25]. Il a été suggéré que les corps huileux pourraient avoir une fonction dissuasive contre les herbivores ou pourraient protéger des températures froides ou des rayons ultraviolets nocifs[22],[23].
Sporophyte
modifierLe sporophyte est constitué d'une tige non ramifiée appelée soie, qui porte une capsule de spores terminale appelée sporange[23]. Les sporanges de Conocephalum sont portés sous des structures gamétophytes pédonculées appelées archégoniophores[4]. Contrairement aux mousses, le sporophyte mûrit avant que la soie ne s'allonge[23],[26]. Contrairement aux mousses, les sporophytes de l'hépatique n'ont pas de stomates, de columelle et de dents péristomiales[22],[23].
Cycle de vie
modifier
Le cycle de vie des hépatiques implique une alternance de générations de gamétophytes haploïdes et de sporophytes diploïdes. La génération de gamétophytes est plus dominante, tandis que la génération de sporophytes est relativement courte. Le gamétophyte produit des gamètes haploïdes, des ovules et des spermatozoïdes, qui fusionnent pour former un zygote diploïde. Le zygote se développe ensuite en sporophyte qui produit finalement des spores haploïdes par méiose. Le sporophyte a besoin de nutriments fournis par le gamétophyte pour soutenir sa croissance et son développement.
Le cycle de vie des hépatiques Marchantia s'applique également à Conocephalum, à l'exception du fait que Conocephalum n'a pas d'anthéridiophore pédonculé et a plutôt de petites têtes anthéridiennes plates à la surface du thalle[23].
Reproduction
modifier_1912.jpg)
Les hépatiques se reproduisent par reproduction sexuée et asexuée[5]. Dans les populations naturelles, la forte variation génétique observée suggère que la reproduction sexuée pourrait dominer. Les espèces de Conocephalum sont dioïques[9], ce qui signifie que les structures reproductrices mâles et femelles sont produites sur des plantes séparées[27].
Reproduction sexuée
modifierChez Conocephalum, les organes reproducteurs mâles et femelles sont intégrés dans des réceptacles sur des plantes séparées. Sur les plantes mâles, le réceptacle est légèrement surélevé, dépourvu de tige, et souvent de forme circulaire ou ovale. Les anthéridies sont intégrées dans le réceptacle[5] et à maturité, le sperme est libéré dans l'air[5],[23],[28]. En revanche, sur les plantes femelles, les réceptacles sont en forme de dôme, avec plusieurs lobes tombants à l'extrémité d'une tige dressée. Les réceptacles sont souvent décrits comme un minuscule parapluie, avec les archégones en dessous[5].
Les gamétophytes produisent des œufs et des spermatozoïdes dans les archégones et les anthéridies, respectivement[20]. La fécondation se produit lorsque le sperme atteint l'œuf dans l'archégone d'une plante femelle[21]. Une fois la fécondation effectuée, l'ovule dans un archégone se développe en sporophyte[20]. Les sporanges matures situés sous le réceptacle ressemblent à des capsules noires. Ces capsules s'ouvrent pour libérer à la fois des spores et des élatères[21], qui sont dispersés principalement par le vent[20],[21]. Les élatères ont pour fonction de propulser les spores pendant la dispersion[26].
Les élatères de Conocephalum sont uniques et présentent une grande variabilité de forme, de taille et de nombre. Souvent, l'abondance des élatères dans une capsule est 2 à 3 fois plus abondante que celle des spores[29]. Les élatères se forment à partir d'une cellule mère initiale qui se développe en une cellule diploïde avec des épaississements en spirale. En revanche, les spores se développent à partir d'une cellule mère diploïde initiale qui forme finalement des spores haploïdes par méiose[22].
_1022.jpg)
Reproduction asexuée
modifierLa production de gemmes est une méthode courante de reproduction asexuée chez les hépatiques[5],[26],[30]. Les gemmes sont de petits paquets de tissus constitués de cellules haploïdes génétiquement identiques à celles de la plante mère. Elles sont dispersées par les précipitations et finissent par se développer en de nouveaux individus[30]. Chez C. conicum, les gemmes sont situées sur les couches inférieures du thalle et sont libérées lorsque le thalle se dégrade[4]. En revanche, C. salebrosum ne produit pas de gemmes.
La reproduction végétative peut se produire lorsqu'un morceau du thalle se détache et est transporté loin de la plante mère[5]. Les individus résultant de la reproduction végétative sont génétiquement identiques à la plante mère et, par conséquent, les colonies clonales existent souvent en tant que mâles ou femelles[26].
Les espèces de Conocephalum sont pérennes, ce qui signifie qu'elles peuvent hiverner et produire de nouvelles pousses au printemps. Ces nouveaux bourgeons sont recouverts et protégés par de petites écailles[9].
Biochimie
modifier
De nombreuses hépatiques produisent différents terpènes et composés aromatiques[10],[11]. Les terpénoïdes et les composés aromatiques sont souvent accumulés dans les corps huileux de nombreuses hépatiques, y compris Conocephalum. Au sein de Conocephalum, une variation considérable des espèces a été identifiée en fonction de la composition chimique et ces composés ont été utilisés pour identifier différentes espèces cryptiques[12].
Trois groupes différents de Conocephalum ont été identifiés en fonction de leurs composés volatils primaires uniques. Par exemple, le composé cubébol, un alcool sesquiterpénique, est caractéristique de C. salebrosum[12]. Il a également été noté que C. supradecompositum a une composition chimique distincte par rapport à C. conicum également, principalement que la teneur en monoterpénoïdes de C. supradecompositum est bien inférieure à celle observée chez C. conicum[18].
Un alcool sesquiterpénique unique connu sous le nom de conocephalenol a été identifié et extrait de C. conicum[10],[11]. Le conocephalenol possède un squelette chimique unique qui est caractéristique d'un sesquiterpène présent dans les algues rouges[31].
Associations avec d'autres espèces
modifierInteractions fongiques
modifierConocephalum peut former des associations avec des champignons qui sont similaires aux associations mycorhiziennes observées chez les plantes vasculaires. Des analyses moléculaires ont démontré que Conocephalum contenait des endophytes fongiques du groupe de champignons connus sous le nom de Glomeromycota[32].
Conocephalum conicum colonise souvent les sols nus ou les substrats rocheux, où les nutriments minéraux peuvent souvent être limitants. L'endophyte fongique établit une relation complexe avec C. conicum, caractérisée par la formation d'arbuscules. Ces champignons forment un mycélium très ramifié à l'extérieur de la plante qui colonise ensuite l'extérieur des rhizoïdes et passe dans le gamétophyte. L'infection fongique induit la croissance d'hyphes fongiques dans les cellules hôtes de C. conicum. Cette association de l'hyphe fongique avec les plastes de l'hôte suggère que la photosynthèse produite par C. conicum est probablement transférée au champignon. Une situation similaire concernant cette association fongique a également été identifiée chez l'hépatique thalloïde Pellia epiphylla. Bien que ces associations soient courantes chez les plantes vasculaires, elles ont rarement été décrites chez les plantes non vasculaires[33].
Interactions avec les animaux
modifierHerbivorie
modifierEnviron 25 espèces de papillons de nuit endémiques à l'Asie de l'Est s'associent exclusivement à Conocephalum[9]. Le stade larvaire des papillons Epimartyria pardella se nourrit de C. conicum[34],[35]. De plus, l'espèce fongique Loreleia marchantiae se nourrit également de C. conicum[9].
Pathogènes
modifierLe pathogène fongique appartenant au genre Pythium a souvent été isolé à partir de rhizoïdes et de thalles infectés de Conocephalum[9]. Bryoscyphus conocephali est un autre pathogène fongique qui a été associé à C. conicum[36].
Applications humaines
modifierConocephalum comme bioindicateur de pollution
modifierConocephalum conicum a été identifié comme étant tolérant aux métaux lourds et a donc été suggéré comme ayant un rôle possible comme bioindicateur de pollution. Conocephalum conicum absorbe les ions à la fois du sol et de l'atmosphère. Par conséquent, la contamination par les métaux lourds de C. conicum est liée non seulement à la pollution de l'air, mais aussi à la contamination environnementale provenant de différentes sources[37]. Des recherches récentes ont également examiné C. conicum comme bioindicateur de pollution au cadmium. Le cadmium est un métal toxique et considéré comme le troisième contaminant le plus important, après le mercure et le plomb. Il a été démontré que Conocephalum conicum réagit au stress du cadmium en modifiant son activité biologique. Ces changements biologiques pourraient être utilisés comme biomarqueurs de la pollution au cadmium[38].
-Aflatoxin_B1_Structural_Formulae_V.1.svg)
Activité antifongique
modifierIl a été suggéré que Conocephalum conicum avait un rôle possible dans la gestion des maladies d'origine alimentaire causées par des espèces de champignons Aspergillus. Aspergillus produit des toxines très puissantes, des cancérogènes, appelés aflatoxines. Les aflatoxines sont nocives pour les plantes et les animaux. Aspergillus peut provoquer des maladies dans de nombreuses cultures importantes, qui peuvent à terme provoquer des maladies chez l'homme. Il a été démontré que Conocephalum possède une variété de composés bioactifs qui favorisent la propriété antifongique contre Aspergillus[24].
Ethnomédecine
modifierPartout en Amérique du Nord, en Chine et en Inde, des hépatiques telles que Conocephalum ont été utilisées à des fins ethnomédicales. Conocephalum est connu pour être important pour les tribus Bhotia, Raji, Tharus et Boxas dans le district de Pithoragarh de Kumaon Himalaya. Conocephalum conicum est utilisé pour traiter les brûlures, et l'extrait de C. conicum est également utilisé dans le traitement des calculs biliaires. Conocephalum a également démontré une activité antidote contre les morsures de serpents venimeux. Le rôle de Conocephalum dans la médecine moderne n'a pas encore été étudié[24].
Industrie cosmétique
modifierLes hépatiques contiennent souvent des composés très piquants[39]. Les espèces de Conocephalum dégagent souvent une odeur caractéristique de la térébenthine. On pense que cette odeur est liée à la présence de monoterpénoïdes. Le composé conocephalenol est largement utilisé dans l'industrie cosmétique pour ses propriétés odorantes[31].
Galerie de photos
modifier-
 Conocephalum conicum, le thalle est nettement brillant
Conocephalum conicum, le thalle est nettement brillant -
 Conocephalum salebrosum, le thalle est d'apparence terne
Conocephalum salebrosum, le thalle est d'apparence terne -
 Conocephalum conicum, thalle
Conocephalum conicum, thalle -
 Conocephalum conicum, pores d'air visibles sur la surface supérieure du thalle
Conocephalum conicum, pores d'air visibles sur la surface supérieure du thalle -
 Conocephalum salebrosum, coupe transversale du thalle montrant les cellules photosynthétiques
Conocephalum salebrosum, coupe transversale du thalle montrant les cellules photosynthétiques -
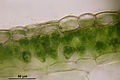 Conocephalum salebrosum, coupe transversale du thalle montrant les cellules photosynthétiques
Conocephalum salebrosum, coupe transversale du thalle montrant les cellules photosynthétiques -
 Conocephalum conicum, pore d'air présent sur la surface supérieure du thalle
Conocephalum conicum, pore d'air présent sur la surface supérieure du thalle -
 Conocephalum conicum, sporophytes (noirs) suspendus sous les archégoniophores pédonculés en forme de parapluie
Conocephalum conicum, sporophytes (noirs) suspendus sous les archégoniophores pédonculés en forme de parapluie -
 Conocephalum salebrosum, sporophytes (noirs) suspendus sous les archégoniophores pédonculés en forme de parapluie
Conocephalum salebrosum, sporophytes (noirs) suspendus sous les archégoniophores pédonculés en forme de parapluie -
 Conocephalum conicum, sporophytes (noirs) suspendus sous les archégoniophores
Conocephalum conicum, sporophytes (noirs) suspendus sous les archégoniophores -
 Conocephalum conicum, anthéridie
Conocephalum conicum, anthéridie -
 Conocephalum salebrosum, spores et élatères
Conocephalum salebrosum, spores et élatères -
 Conocephalum salebrosum, spores et élatères
Conocephalum salebrosum, spores et élatères -
 Conocephalum salebrosum, pore infrarouge présent sur la surface supérieure du thalle
Conocephalum salebrosum, pore infrarouge présent sur la surface supérieure du thalle
.jpg)
_7808.JPG)
_7810.JPG)
_4442.JPG)

_4502.JPG)
_4504.JPG)
_4496.JPG)
Liens externes
modifier
- Ressources relatives au vivant :
- Australian Plant Name Index
- Dyntaxa
- EPPO Global Database
- Flora of North America
- Global Biodiversity Information Facility
- iNaturalist
- Index Hepaticarum
- Interim Register of Marine and Nonmarine Genera
- Nálezová databáze ochrany přírody
- NBN Atlas
- NDFF Verspreidingsatlas
- Nederlands Soortenregister
- PLANTS Database
- Système d'information taxonomique intégré
- Tropicos
- Notice dans un dictionnaire ou une encyclopédie généraliste :
Références
modifier- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Conocephalum » (voir la liste des auteurs).
- (en) J. Szweykowski, K. Buczkowska et I. J. Odrzykoski, « Conocephalum salebrosum (Marchantiopsida, Conocephalaceae) – a new Holarctic liverwort species », Plant Systematics and Evolution, vol. 253, no 1, , p. 133–158 (ISSN 1615-6110, DOI 10.1007/s00606-005-0301-0, S2CID 20076390, lire en ligne)
- AKIYAMA, H. (2022). Morphological and ecological diversification of Conocephalum conicum complex in Japan and Taiwan. Humans Nat, 32, 1-45.
- « Vascular Plants of the Gila Wilderness-- Conocephalum salebrosum », sur wnmu.edu (consulté le )
- (en-US) « Conocephalum conicum | Introduction to Bryophytes » (consulté le )
- « Minnesota Seasons - snakeskin liverwort », sur minnesotaseasons.com (consulté le )
- (en) Chi Zhang, Xinlu Chen, Barbara Crandall-Stotler, Ping Qian, Tobias G. Köllner, Hong Guo et Feng Chen, « Biosynthesis of methyl (E)-cinnamate in the liverwort Conocephalum salebrosum and evolution of cinnamic acid methyltransferase », Phytochemistry, vol. 164, , p. 50–59 (ISSN 0031-9422, PMID 31078779, DOI 10.1016/j.phytochem.2019.04.013, S2CID 153298906, lire en ligne)
- « NatureServe Explorer 2.0 », sur explorer.natureserve.org (consulté le )
- « Species Summary », sur a100.gov.bc.ca (consulté le )
- « Bryophyte Ecology Volume 4 ebook | Bryophyte Ecology | Michigan Technological University », sur digitalcommons.mtu.edu (consulté le )
- (en) Motoo Tori, Masakazu Sono, Katsuyuki Nakashima, Yoko Nakaki et Yoshinori Asakawa, « Synthesis and relative structure of conocephalenol, a sesquiterpene alcohol isolated from the european liverwort Conocephalum conicum », Journal of the Chemical Society, Perkin Transactions 1, no 2, , p. 447–450 (ISSN 1364-5463, DOI 10.1039/P19910000447, lire en ligne)
- (en) Motoo Tori, Katsuyuki Nakashima, Yoshinori Asakawa, Joseph D. Connolly, Leslie J. Harrison, David S. Rycroft, Jagdemba Singh et Neill Woods, « Structure of conocephalenol, a sesquiterpenoid alcohol from the European liverwort Conocephalum conicum: determination of the absolute configuration by total synthesis », Journal of the Chemical Society, Perkin Transactions 1, no 5, , p. 593–597 (ISSN 1364-5463, DOI 10.1039/P19950000593, lire en ligne)
- (en) Agnieszka Ludwiczuk, Ireneusz J. Odrzykoski et Yoshinori Asakawa, « Identification of cryptic species within liverwort Conocephalum conicum based on the volatile components », Phytochemistry, vol. 95, , p. 234–241 (ISSN 0031-9422, PMID 23835199, DOI 10.1016/j.phytochem.2013.06.011, lire en ligne)
- Raymond E. Stotler et Barbara Crandall-Stotler, « A Synopsis of the Liverwort Flora of North America North of Mexico », Annals of the Missouri Botanical Garden, vol. 102, no 4, , p. 574–709 (ISSN 0026-6493, DOI 10.3417/2016027, S2CID 89829947, lire en ligne)
- ↑ H Ki, Eiji Nitasaka, Ireneusz Odrzykoski et T Yamazak, « Phylogenetic relationships among taxa of the liverwort Conocephalum conicum (Conocephalaceae) revealed by psbA sequence », Genes & Genetic Systems, vol. 76, no 5, , p. 279–88 (PMID 11817643, DOI 10.1266/ggs.76.279
 , lire en ligne)
, lire en ligne)
- C Sérgio, C Vieira, D Claro et C Garcia, « Conocephalum salebrosum Szweykowski, Buczkowska & Odrzykoski (Marchantiopsida): modelling the occurrence of a hygrophytic species new to the bryoflora of Portugal, Madeira and Azores », Journal of Bryology, vol. 33, no 1, , p. 30–34 (ISSN 0373-6687, DOI 10.1179/1743282010Y.0000000004, S2CID 85286389, lire en ligne)
- ↑ « NatureServe Explorer 2.0 », sur explorer.natureserve.org (consulté le )
- ↑ (en) Ireneusz J. Odrzykoski et Jerzy Szweykowski, « Genetic differentiation without concordant morphological divergence in the thallose liverwortConocephalum conicum », Plant Systematics and Evolution, vol. 178, no 3, , p. 135–152 (ISSN 1615-6110, DOI 10.1007/BF00937961, S2CID 32759989, lire en ligne)
- (en) Yoshinori Asakawa, Reiko Matsuda et Reiji Takeda, « Mono- and sesquiterpenoids of Conocephalum supradecompositum », Phytochemistry, vol. 20, no 6, , p. 1423–1424 (ISSN 0031-9422, DOI 10.1016/0031-9422(81)80055-6, lire en ligne)
- Cros, R. M., & Buczkowska, K. (2009). Conocephalum salebrosum (Marchantiopsida) new to Spain. Cryptogamie, 30(1), 203.
- « About Mosses and Liverworts », sur www.illinoiswildflowers.info (consulté le )
- « Snakeskin Liverwort (Conocephalum salebrosum) », sur www.illinoiswildflowers.info (consulté le )
- (en-US) « Class Marchantiopsida | Introduction to Bryophytes » (consulté le )
- « Bryophyte Ecology Volume 1 ebook | Bryophyte Ecology | Michigan Technological University », sur digitalcommons.mtu.edu (consulté le )
- (en) K. Negi, A. K. Asthana et P. Chaturvedi, « GC–MS analysis and antifungal activity of acetone extract of Conocephalum conicum (L) Underw (Liverwort) against aflatoxins producing fungi », South African Journal of Botany, vol. 131, , p. 384–390 (ISSN 0254-6299, DOI 10.1016/j.sajb.2020.02.035 , S2CID 216325490, lire en ligne)
- Xiaolan He, Yu Sun et Rui-Liang Zhu, « The Oil Bodies of Liverworts: Unique and Important Organelles in Land Plants », Critical Reviews in Plant Sciences, vol. 32, no 5, , p. 293–302 (ISSN 0735-2689, DOI 10.1080/07352689.2013.765765, S2CID 55444410, lire en ligne)
- « Biology 321 - UBC », sur www3.botany.ubc.ca (consulté le )
- ↑ Juan Carlos Villarreal et Susanne S. Renner, « Correlates of monoicy and dioicy in hornworts, the apparent sister group to vascular plants », BMC Evolutionary Biology, vol. 13, no 1, , p. 239 (ISSN 1471-2148, PMID 24180692, PMCID 4228369, DOI 10.1186/1471-2148-13-239 )
- ↑ Masaki Shimamura, Tomio Yamaguchi et Hironori Deguchi, « Airborne sperm of Conocephalum conicum (Conocephalaceae) », Journal of Plant Research, vol. 121, no 1, , p. 69–71 (PMID 18058191, DOI 10.1007/s10265-007-0128-6, S2CID 37902329)
- ↑ Naofumi Kitagawa, « A study of Conocephalum supradecompositum, Marchantiales, Hepaticae », Acta Phytotaxonomica et Geobotanica, vol. 33, , p. 179–189 (DOI 10.18942/bunruichiri.KJ00001079155, lire en ligne)
- AJE Smith, The Liverworts of Britain and Ireland, Cambridge University Press, Cambridge,
- Anaïs Pannequin, Erik Laurini, Laurent Giordano, Alain Muselli, Sabrina Pricl et Aura Tintaru, « Caution: Chemical Instability of Natural Biomolecules During Routine Analysis », Molecules, vol. 25, no 14, , p. 3292 (ISSN 1420-3049, PMID 32698478, PMCID 7397321, DOI 10.3390/molecules25143292 )
- ↑ R. Ligrone, A. Carafa, E. Lumini, V. Bianciotto, P. Bonfante et J. G. Duckett, « Glomeromycotean associations in liverworts: a molecular, cellular, and taxonomic analysis », American Journal of Botany, vol. 94, no 11, , p. 1756–1777 (ISSN 0002-9122, PMID 21636371, DOI 10.3732/ajb.94.11.1756, lire en ligne)
- ↑ (en) R. Ligrone et C. Lopes, « Cytology and development of a mycorrhiza-like infection in the gametophyte of Conocephalum conicum (L.) Dum. (Marchantiales, Hepatophyta) », New Phytologist, vol. 111, no 3, , p. 423–433 (ISSN 1469-8137, PMID 33874002, DOI 10.1111/j.1469-8137.1989.tb00705.x , lire en ligne)
- ↑ « Conocephalum conicum | NBN Atlas », sur species.nbnatlas.org (consulté le )
- ↑ Donald R. Davis et Jean-François Landry, « A review of the North American genus Epimartyria (Lepidoptera, Micropterigidae) with a discussion of the larval plastron », ZooKeys, no 183, , p. 37–83 (ISSN 1313-2989, PMID 22573948, PMCID 3332027, DOI 10.3897/zookeys.183.2556 )
- ↑ « Conocephalum conicum (L.) Dumort. - Encyclopedia of Life », sur eol.org (consulté le )
- ↑ (en) Adriana Basile, Sergio Sorbo, Barbara Conte, Manuela Cardi et Sergio Esposito, « Ultrastructural changes and Heat Shock Proteins 70 induced by atmospheric pollution are similar to the effects observed under in vitro heavy metals stress in Conocephalum conicum (Marchantiales – Bryophyta) », Environmental Pollution, vol. 182, , p. 209–216 (ISSN 0269-7491, PMID 23933125, DOI 10.1016/j.envpol.2013.07.014, lire en ligne)
- ↑ Viviana Maresca, Gennaro Lettieri, Sergio Sorbo, Marina Piscopo et Adriana Basile, « Biological Responses to Cadmium Stress in Liverwort Conocephalum conicum (Marchantiales) », International Journal of Molecular Sciences, vol. 21, no 18, , p. 6485 (ISSN 1422-0067, PMID 32899890, PMCID 7555243, DOI 10.3390/ijms21186485 )
- ↑ (en) Yoshinori Asakawa, Reiko Matsuda et Tsunematsu Takemoto, « Mono- and sesquiterpenoids from Wiesnerella denudata », Phytochemistry, vol. 19, no 4, , p. 567–569 (ISSN 0031-9422, DOI 10.1016/0031-9422(80)87016-6, lire en ligne)