Arenicola marina
L’arénicole, Arenicola marina, souvent appelée « ver de vase » ou « ver noir » est un ver annélide dont le corps est constitué d’une série d’anneaux successifs, appelés segments ou métamères. Chez l’arénicole ces métamères sont subdivisés en 2 à 5 annelets superficiels. Le ver appartient à la classe des annélides polychètes (de poly = plusieurs et chètes = soies) : les métamères de la partie la plus renflée sont munis de pieds ou parapodes garnis de soies plus ou moins rigides, mobiles, qui permettent à l’animal de prendre appui sur le support pour s’immobiliser ou se déplacer. À l'âge adulte, sa longueur moyenne est d'une quinzaine de centimètres, avec un maximum d'une trentaine de centimètres[1].

| Règne | Animalia |
|---|---|
| Embranchement | Annelida |
| Classe | Polychaeta |
| Ordre | Capitellida |
| Famille | Arenicolidae |
| Genre | Arenicola |
Noms régionaux
modifierBien connu des pêcheurs qui le désignent de plusieurs autres noms locaux comme chique, buzuc , buzin, etc. Il existe aussi un mot spécifique à la Normandie occidentale (Calvados, Manche) sando[n] ou sandron issu de l'ancien norois *sandormr, de sandr « sable » et ormr « serpent » cf. danois sandorm, même sens.
Morphologie
modifierLe corps est approximativement cylindrique, renflé dans sa région antérieure très déformable, plus étroit dans sa région postérieure[2]. On peut y distinguer, de manière quelque peu conventionnelle, quatre parties :
La tête
modifierConstituée par :
- Le prostomium. C’est un petit bourrelet triangulaire dorsal, à base tournée vers l’avant, placé en arrière de la bouche. Sa surface est divisée en trois lobes par deux sillons disposés en Y et ne laisse apparaître par ailleurs aucun organe particulier. Il peut être rétracté dans une poche, la poche nuchale, et donc ne pas être immédiatement visible.
- Le péristomium, également appelé segment buccal, qui porte la bouche, munie d’une trompe exsertile (trompe pharyngienne qui s'évagine), globuleuse, couverte de nombreuses papilles. Ce segment correspond aux deux premiers annelets, pas toujours nettement séparés, de la région antérieure.
- Un segment sans soies (« achète ») représenté par les deux annelets suivants.
Le thorax
modifierLe thorax est composé de six métamères munis de soies (= sétigères). Le premier est formé de deux annelets, le deuxième de trois, le 3e de quatre, le 5e et le 6e de cinq. Les parapodes sont situés sur l’avant-dernier annelet de chaque métamère, plus saillant que les autres, chacun d’entre eux est formé d’une rame dorsale ou notopode (de noto = dos et pode = pied) garni de longues soies épineuses et d’une rame ventrale ou neuropode (située du côté du cordon nerveux, qui est ventral). Cette dernière est équipée de soies courtes, en forme de « S », les uncinis (= crochets).
L’abdomen
modifierL’abdomen est formé par les 13 métamères suivants, semblables aux derniers du thorax, mais munis de branchies, ramifiées, contractiles, rouges, situées près de la rame dorsale des parapodes.
Wells[3] réunit le thorax et l’abdomen sous l’appellation de « tronc » comportant donc 19 métamères munis de parapodes (« sétigères »).
La queue
modifierNettement plus étroite que la région précédente, la queue est constituée d’un nombre indéterminé de métamères, sans parapodes ni branchies, chacun d’entre eux représenté par un annelet. Les métamères, très courts dans la partie basale, sont allongés dans la partie distale dont l’extrémité porte l’anus. La peau de l’arénicole sécrète une substance fluorescente gris-vert, quelque peu irritante, qui colore les doigts lorsqu’on la manipule.
Éléments d’anatomie
modifier- Tube digestif : comporte la trompe, suivie d’un œsophage muni de deux diverticules, les glandes œsophagiennes, puis vient l’estomac renflé, recouvert d’un tissu jaune, le tissu chloragogène (tissu qui accumule les déchets mais sert probablement aussi de réserve) et enfin un long intestin terminé par l’anus.
- Appareil circulatoire : le sang est contenu dans un système complexe de vaisseaux clos ; il est coloré en rouge par l'érythrocruorine dissoute dans le plasma et mis en mouvement par deux cœurs latéraux (1 oreillette et 1 ventricule chacun) ainsi que par les vaisseaux à parois contractiles.
- Appareils excréteur et génital : six paires de néphridies (3 thoraciques + 3 abdominales) constituées d’un pavillon à épithélium vibratile, un sac néphridien et une vésicule contractile qui s’ouvre par un pore allongé, en arrière de la limite supérieure des neuropodes. Les gonades sont situées à l’extrémité postérieure du pavillon des 5 dernières néphridies, qui assurent l’évacuation des produits génitaux, et sont souvent masquées par la vésicule.
- Système nerveux : des ganglions cérébroïdes aux contours mal définis sont plaqués contre le « plafond » du prostomium, ils sont suivis par un collier péri-œsophagien (= connectif) puis un cordon nerveux longitudinal, dépourvu de ganglions, dont la présence est révélée, extérieurement, par une ligne blanchâtre, au fond d’un sillon, à la face ventrale de l’animal. Dans le segment buccal se trouvent deux statocystes jaunes. Ces organes sensoriels permettent à l’arénicole de connaître sa position dans l’espace (savoir si elle est en position horizontale ou verticale, si elle a la tête tournée vers le haut ou vers le bas etc.)
Biologie
modifierLa vie dans le sédiment
modifier
L’arénicole se loge entre 10 et 20, voire 40 cm de profondeur dans le sédiment sableux ou sablo-vaseux. Il est généralement absent des sédiments dont la taille moyenne des particules est inférieure à 80 μm et son abondance diminue lorsque cette taille dépasse 200 μm. Le ver aménage un système en forme de U, constitué de deux parties[5].
La galerie
modifierLa galerie est un tube, approximativement en forme de L ou de J (selon la manière dont on regarde le tube) qui occupe la branche horizontale et une branche verticale du U. Elle est gainée d’un sédiment de couleur claire (gris ou rouille, ce qui dénote son état oxydé) consolidé par du mucus sécrété par l’animal. La branche verticale comporte, dans sa partie haute, une portion plus étroite, la « cheminée » caudale, dans laquelle seule pénètre la queue du ver, notamment lorsqu’il vient déféquer. Les excréments, constitués essentiellement de sédiment, forment un turricule (en forme de tortillon) ou un cône plus ou moins élevé au-dessus de la surface voisine. L’arénicole occupe généralement la partie la plus large de la galerie, c’est-à-dire la branche horizontale et la partie inférieure de la branche verticale.
La partie antérieure
modifierElle est principalement représentée par la deuxième branche ascendante du U. Elle est très variable dans sa forme et sa structure, selon la nature du sédiment et les activités de l’arénicole. C’est un espace vaguement conique auquel correspond en surface une zone déprimée, voire un trou menant à un puits mais généralement il est comblé par du sédiment fortement remanié par l’animal, qui y fait des incursions sporadiques, et par le courant d’eau qu’il y entretient.
L’arénicole provoque dans ce système deux courants de sens opposés : Un courant d’eau provoqué par des ondes de dilatations et de contractions qui parcourent le corps de l’arrière vers l’avant et font circuler l’eau de la queue vers la tête en passant essentiellement par la face dorsale du ver. Le flux engendré mesure de 1.2 à 7.2 ml par minute pour un animal de 17.2 g[6], 1,5 ml par minute pour une arénicole « standard » (poids sec = 0.5 g) parcourue par 7 contractions à la minute à 15 °C[7]. Cette circulation d’eau permet à l’animal de respirer et l’oxygène qu’elle transporte est aussi responsable de la couleur claire du sédiment à la périphérie de la galerie. Par ailleurs, ce courant d’eau percole à travers le sédiment de la partie antérieure du système dont il modifie la composition et la stabilité (la pression engendrée atteint, au moins temporairement, 20 cm d’eau[7]). Chaque cycle d’irrigation comporte trois phases : a, un déplacement du corps vers l’arrière, b, pompage et déplacement du corps vers l’avant, c, pompage vers l’arrière. La phase b est la plus importante[8]. Une circulation de sédiment provoquée par le fait que l’animal avale du sable (particules < 2 mm) et de la vase à la base de la région antérieure et les rejette au-dessus de l’orifice postérieur. Faisant partie des organismes marins psammivores (de psammivore, littéralement « mangeur de sable », issu du grec ancien psámmos et du latin vore). Le ver se nourrit des détritus organiques ainsi que de la microflore et de la microfaune contenus dans le sédiment. Une partie de ces éléments provient du dépôt qui se forme dans la dépression superficielle au-dessus de la branche antérieure du U (d’où le qualificatif de déposivore = « deposit-feeder », attribué à l’arénicole[6] On a par ailleurs avancé (Krüger, 1959) que le courant d’eau postéro-antérieur était susceptible d’enrichir le sédiment des éléments organiques qu’il transporte contribuant ainsi à l’alimentation de l’arénicole. Cette hypothèse ne semble pas devoir être retenue[7].
Ces activités se déroulent lorsque le biotope de l’animal est immergé. Les cycles de « ventilation » de la galerie durent de 10 à 15 minutes[6] ils sont séparés par des périodes d’alimentation (durée environ 7 minutes[9]), ou de repos. Ces cycles sont commandés par le système nerveux : un pacemaker dans la chaîne ventrale pour la ventilation, un autre dans la paroi œsophagienne pour l’alimentation, ils ont un caractère automatique qui semble indépendant des conditions extérieures[9].
À basse mer, selon les conditions de drainage du sédiment, la galerie peut se vider, le ver se tient alors dans la partie postérieure verticale et respire l’oxygène de l’air dans une ambiance saturée d’humidité, éventuellement, il entraîne des bulles d’air jusqu’au contact des branchies[5]. Si la galerie reste en eau, l’animal peut épuiser l’oxygène dissous et être confronté à des concentrations élevées d’hydrogène sulfuré (H2S, HS‾), il se met alors en métabolisme anaérobie ; lorsqu’il est de nouveau en immersion, il oxyde l’hydrogène sulfuré accumulé dans le liquide cœlomique en thiosulfate. Il se remet de ce stress en moins d’une heure[10]
Reproduction
modifierLes sexes sont séparés et non identifiables extérieurement. Il y aurait en moyenne 3,75 femelles pour 1 mâle[11],[12]. Les gonades, très petites, libèrent les éléments génitaux dans le cœlome où ils poursuivent leur évolution. Les ovocytes sont séparés les uns des autres et aisément reconnaissables. Les spermatozoïdes (spermatocytes en réalité) sont réunis en masses arrondies ayant l’aspect d’une mûre (« morula ») qui se dissocient avant leur émission sous l’effet d’un « facteur de maturation » sécrété par le prostomium[13].
Le pic de reproduction a lieu en automne (octobre-novembre). Les gamètes sont émis simultanément, sur une courte période de 4-5 jours lors de basses mers de vive eau. Ce synchronisme est dû à des signaux chimiques (phéromones) libérés par les individus reproducteurs[14]. Les ovocytes restent dans la galerie, les spermatozoïdes s’étalent en galettes à la surface du sédiment et sont entraînés dans les galeries, où a lieu la fécondation, par le courant d’eau engendré par les femelles lorsque la mer remonte.
Les larves se développent dans la galerie de la femelle. Les post-larves rampent dans la couche superficielle du sédiment et peuvent être entraînées par les courants mais ne mènent pas une vie franchement planctonique. Les juvéniles s’installent sur des territoires différents de ceux occupés par les individus âgés[15]. Les individus peuvent atteindre la maturité sexuelle au bout d’un an et plus généralement au cours de leur deuxième année. La durée de vie serait d’au moins 5-6 ans. La mortalité des adultes atteindrait 40 % après la reproduction selon Newell (1948), d’autres auteurs indiquent une mortalité annuelle moyenne de 22 %.
Importance écologique et économique
modifier
Les populations d’arénicoles peuvent atteindre des densités de 100-150 individus par mètre carré et constituer 30 % de la biomasse d’une plage ordinaire[16]. Leur activité de fouissage, de pompage et de prise de nourriture peut remanier radicalement le sédiment (phénomène appelé bioturbation) modifier sa structure (les particules fines < 2 mm, sont ramenées à la surface, les éléments grossiers restent en profondeur) ainsi que la circulation et les caractéristiques physico-chimiques de l’eau qui y est contenue (notamment sa concentration en oxygène). Cette espèce peut donc imprimer une marque très forte aux estrans qu’elle habite.

C’est une source de nourriture importante pour des poissons plats et plusieurs espèces d’oiseaux limicoles. Généralement le prédateur ne parvient à prélever que la portion terminale de la queue. Celle-ci retrouve sa longueur initiale par extension des métamères restants. Ces amputations sont donc subies sans grand dommage pour l’arénicole, la croissance s’arrête cependant lorsqu’elles interviennent au rythme de une fois par semaine[17].
Les arénicoles sont des appâts très recherchés pour la pêche des poissons marins, elles font l’objet d’un commerce florissant. Leur récolte s’effectue par bêchage du sédiment, extraction de l’animal grâce à une pompe(dans le sable) voire (aux Pays-Bas) à l’aide de bateaux spécialement équipés. On a calculé qu’aux Pays-Bas, 29 à 36 millions de vers sont débarqués chaque année[18]. Le bêchage à grande échelle (plusieurs centaines de mètres carrés) peut avoir des effets préoccupants sur la faune, voire la flore, des estrans concernés, même si les populations d’arénicoles se reconstituent en quelques mois par émigration d’individus (notamment de juvéniles) à partir des régions voisines.
L’arénicole est par ailleurs utilisée dans les bioessais concernant les effets de toxiques dans le sédiment et dans diverses études concernant des phénomènes comme la respiration par exemple. On a suggéré le recours à son hémoglobine comme substitut du sang dans les transfusions pour l’homme[19].
Espèces voisines
modifierParmi les cinq espèces d’Arenicola mentionnées par Fauvel [réf. souhaitée], celles qui sont dépourvues de queue (A. ecaudata et A. grubii) sont actuellement classées dans le genre Arenicolides[20]. Quant à A. claparedii, elle est classée dans le genre Abarenicola. Seules subsistent donc dans le genre Arenicola, A. marina et A. cristata, signalée en Méditerranée (Naples) et abondante sur les côtes de l’Amérique du Nord (Atlantique et Pacifique) et de l’Océan indien tropical. Dans les eaux européennes, une nouvelle espèce, Arenicola defodiens (signalée en note par Fauvel), a été séparée de A. marina[21]. Il atteint une plus grande taille que A. marina, se trouve sur des plages plus exposées et à un niveau inférieur de l’estran[22]. A. defodiens ne semble pas avoir été signalée sur les rivages de l’Europe continentale, elle n’apparaît pas dans la liste des espèces présentes en Manche[23].
Utilisations médicales actuelles ou potentielles
modifierLa molécule utilisée par ce ver pour fixer l'oxygène dans son sang est 250 fois plus petite qu'un globule rouge humain, mais 40 fois plus oxygénante, ce qui présente plusieurs intérêts pour la médecine[24]. Elle pourrait révolutionner le sang en poudre car le leur est lyophillisable, contrairement aux solutions actuelles (développées à partir de cellules humaines ou bovines) qui présentent de nombreux effets secondaires.
La société Hemarina, basée à Morlaix, a développé et mis sur le marché un transporteur d'oxygène universel, dit M101, à partir de l'hémoglobine de l'arénicole[25].
Galerie
modifier-
 Arenicola marina. Les différentes parties du corps.
Arenicola marina. Les différentes parties du corps. -
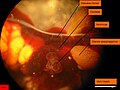 Arenicola marina. Vue du cœur droit. Le ventricule est contracté et presque vide. Les points jaunes sont des ovocytes.
Arenicola marina. Vue du cœur droit. Le ventricule est contracté et presque vide. Les points jaunes sont des ovocytes. -
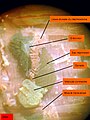 Arenicola marina. Vue d'une néphridie et d'une gonade. Les muscles transversaux qui masquent partiellement ces organes ont été sectionnés
Arenicola marina. Vue d'une néphridie et d'une gonade. Les muscles transversaux qui masquent partiellement ces organes ont été sectionnés -
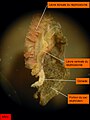 Arenicola marina. Néphrostome vu par la face ventrale et gonade.
Arenicola marina. Néphrostome vu par la face ventrale et gonade. -
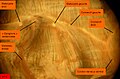 Arenicola marina. Système nerveux. Une incision a été pratiquée du côté droit de la tête, la partie dorsale est rabattue à gauche, le prostomium est vu par la face ventrale. Des faisceaux musculaires recouvrent les éléments nerveux. On distingue au-dessus de l'extrémité du trait de rappel le "X" constitué par la rencontre des deux connectifs et le départ de deux gros cordons, blancs, de fibres nerveuses
Arenicola marina. Système nerveux. Une incision a été pratiquée du côté droit de la tête, la partie dorsale est rabattue à gauche, le prostomium est vu par la face ventrale. Des faisceaux musculaires recouvrent les éléments nerveux. On distingue au-dessus de l'extrémité du trait de rappel le "X" constitué par la rencontre des deux connectifs et le départ de deux gros cordons, blancs, de fibres nerveuses -
 Arenicola marina. Préparation fraîche d'un statocyste vue au microscope.
Arenicola marina. Préparation fraîche d'un statocyste vue au microscope. -
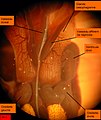 Arenicola marina. Vue des deux cœurs par la face dorsale. Les deux oreillettes ne communiquent pas avec le vaisseau dorsal auquel elles sont accolées. Tissu chloragogène en bas. Les points jaunes épars, sont des ovocytes, libres dans le liquide cœlomique
Arenicola marina. Vue des deux cœurs par la face dorsale. Les deux oreillettes ne communiquent pas avec le vaisseau dorsal auquel elles sont accolées. Tissu chloragogène en bas. Les points jaunes épars, sont des ovocytes, libres dans le liquide cœlomique -
 Arenicola marina. Vue latérale d'un métamère abdominal antérieur
Arenicola marina. Vue latérale d'un métamère abdominal antérieur -
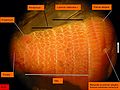 Arenicola marina. Vue latérale de la tête.
Arenicola marina. Vue latérale de la tête.









Références
modifier- Guillaume Fourrier, La pêche à pied : dans votre poche, , 96 p. (ISBN 978-2-85725-916-9, lire en ligne), p. 102.
- Fauvel, P. 1927. Faune de France 16. Polychètes errantes. 494 p Téléchargeable sur :www.faunedefrance.org
- Wells, G.P. 1950. The anatomy of the body wall and appendages in Arenicola marina L., Arenicola claparedii Levinsen and Arenicola ecaudata Johnston. Jour.mar. biol. Ass. U.K. 24 : 1-44.
- Tixier, A. & Gaillard, J.M. 1957. Anatomie animale et dissection. Vigot Frères éditeurs. 375 p.
- (en) Wells, G.P., 1945. The mode of life of Arenicola marina L. Jour. Mar. Biol. Ass. U. K., 26, 170-207
- Toulmond, A. & Dejours, P. 1994. Energetics of the ventilatory piston pump of the lugworm, a deposit–feeding polychaete living in a burrow. Biological Bulletin, 186 : 213-220.
- Riisgård, H.U., Berntsen, I & Tar, B. 1996. The lugworm (Arenicola marina) pump :characteristics, modelling and energy cost. Mar.Ecol. Prog.ser. 138 : 149-156.
- Wells, G.P., 1949a. Respiratory movements of Arenicola marina L.: intermittent irrigation of the tube, and intermittent aerial respiration., Jour. mar. biol. Ass. U. K., 28, 447-478.
- Wells G.P. 1949b. The behaviour of Arenicola marina in sand, and the role of spontaneuos activity cycles. Jour. Mar. biol Ass, UK 29:1-44.
- Völkel, S. Hauschild, K & Grieshaber, M.K. 1995. Sulfide stress and tolerance in the lugworm Arenicola marina during low tide. Mar. Ecol. Prog. Ser. 122 : 205-215.
- Newell, G.E., 1948. A contribution to our knowledge of the life history of Arenicola marina L.., Jour mar. biol. Ass. U.K. 28, 554-580.
- Howie, D.I.D., 1959. The spawning of Arenicola marina (L.). I. The breeding season. Jour. mar. biol. Ass. U.K. 38, 395-406.
- Pacey, A.A. & Bentley, M.G. 1992. The fatty acid 8,11,14-eicosatrienoic acid induces spawning in the male lugworm Arenicola marina. Jour.exp. Biol. 173 : 165-179.
- Hardege, J.D. & Bentley M.G. 1997. Spawning synchrony in Arenicola marina: evidence for sex pheromonal control. Proceedings:Biological Sciences, 264 : 1041-1047
- Hardege, J.D., Bentley, M.G. & L. Snape. 1998. Sediment selection by juvenile Arenicola marina. Mar. Ecol. Prog.Ser. 166 : 187-195
- Arenicola marina : http://zipcodezoo.com/Animals/A/Arenicola_marina/
- Bergman, M.J.N., van der Veer, H.W. & Karczmarski, L. 1988. Impact of tail-nipping on mortality, growth and reproduction of Arenicola marina. Neth. J. Sea Res. 22: 83-90 www.vliz.be/vmdcdata/imis2/imis.php?module=ref&refid=15830
- McLusky, D.S. & Anderson, F.E. 1983. Distribution and population recovery of Arenicola marina and other benthic fauna after bait digging. Mar. Ecol.prog.ser. 11 : 173-179.
- Des chercheurs français inventent le sang de mer www.transfert.net/Des-chercheurs-français-inventent 27k-
- Fauchald, K. 1977. The polychaete worms. Definitions and Keys to the Orders, Families and Genera Natural History Museum of Los Angeles County, Science Series 28, 188p.
- Cadman, P.S. & Nelson-Smith, A. 1993. A new species of lugworm : Arenicola defodiens sp.nov. Jour. mar. biol. Ass.U.K. 73: 213-223
- Everson, C. Two species of lugworm.http://www.glaucus.org.uk/lugs.htm.
- Dauvin, J.C., Dewarumez, J.M. & Gentil, F. 2003. Liste actualisée des espèces d’Annélides Polychètes présentes en Manche. Cah. Bio. Mar. 44 : 67-95.
- Le Pape F, et al.(2015) Advancement in recombinant protein production using a marine oxygen carrier to enhance oxygen transfer in a CHO-S cell line. Artificial Cells, Nanomedicine, and Biotechnology. ;43(3).
- Anaximandre, « HEMOXYCarrier - Français - Transporteur doxygène universel », sur www.hemarina.com (consulté le )
- Les articles des numéros anciens du “Journal of the marine biological Association of the United Kingdom (jmba) sont disponibles sur le site du JMBA

Références taxonomiques
modifier- (en) Référence Catalogue of Life : Arenicola marina (Linnaeus, 1758) (consulté le )
- (fr + en) Référence ITIS : Arenicola marina (Linnaeus, 1758)
- (en) Référence WoRMS : espèce Arenicola marina (Linnaeus, 1758)
Liens externes
modifier- (en) Référence BioLib : Arenicola marina (Linnaeus, 1758)
- (fr) Référence INPN : Arenicola marina (Linnaeus, 1758) (TAXREF)